Original web-page: http://photobiology.info/Zhu.html
ქსინ გუგა ჟუ/Xin-Guang Zhu
მცენარეთა სისტემების ბიოლოგიის ჯგუფი, გამოთვლითი ბიოლოგიის ინსტიტუტი,
ჩინეთის მეცნიერებათა აკადემია/მაქს პლანკის საზოგადოება, Yue Yang Road,
Shanghai, China, 200031
[email protected]
www.picb.ac.cn/~zhuxinguang
შესავალი
წიაღისეული საწვავის შემცირებით და საკვების წარმოებაზე მზარდი მოთხოვნილებით, მცენარეთა ბიოლოგები, ისტორიაში პირველად, მნიშვნელოვან როლს თამაშობენ საზოგადოებისთვის დიდი გამოწვევის გადაწყვეტაში, ანუ, მცენარეთა პროდუქტიულობის გაზრდის ახალი გზების დადგენაში, შეცვლის გლობალურ კლიმატურ პირობებში. ფოტოსინთეზური ენერგიის გადაქცევის ეფექტურობის გაზრდა ჩვენთვის კიდევ ერთი მთავარი გზაა მოსავლის პროდუქტიულობის მკვეთრად გაზრდის მიზნით. სისტემური მოდელების ევოლუციური ალგორითმით გაერთიანება უზრუნველყოფს ერთ ეფექტურ და შესაძლო მექანიზმს ინჟინრის უმაღლესი ფოტოსინთეზისადმი ახალი მიდგომების გამოსავლენად.
ჩვენი ცოდნის უმეტესი ნაწილი ფოტოსინთეზთან დაკავშირებით მიიღება შემცირების შემდგომი მიდგომით, სადაც სისტემის თითოეული ცალკეული კომპონენტი იზოლირებულია და დამოუკიდებლად სწავლობს. ამ მიდგომის შედეგად მივიღეთ დეტალური ბიოფიზიკური და ბიოქიმიური ინფორმაცია ფოტოსინთეზის სხვადასხვა კომპონენტების შესახებ. ამავე დროს, თანამედროვე ბიოტექნოლოგია იძლევა ფოტოსინთეზში თითქმის ყველა გენის მარტივად ან ქვევით რეგულირებას. მიუხედავად ყველა ამ შესაძლებლობისა და ცოდნისა, ჩვენ ჯერ კიდევ შორს ვართ იმის შესაძლებლობისგან, რომ ზუსტად დავაყენოთ ფოტოსინთეტიკური აპარატურა უფრო მაღალი ფოტოსინთეზური მაჩვენებლისთვის. ეს გასაკვირი არ არის იმის გათვალისწინებით, რომ ფოტოსინთეზური ენერგიის გადაქცევის ეფექტურობა არის სისტემის საკუთრება, ანუ ის განისაზღვრება სისტემის სხვადასხვა კომპონენტს შორის ურთიერთქმედებით, რაც არ არის განსაზღვრული სისტემის რომელიმე ინდივიდუალური კომპონენტის მიერ.
იმის გათვალისწინებით, რომ ფოტოსინთეზი არის როგორც საკვების, ისე ენერგიის წარმოების საფუძველი და ჩვენს საზოგადოებაში გაზრდილი მოთხოვნილება, როგორ შეგვიძლია გამოვავლინოთ უფრო მაღალი ფოტოსინთეზი? სხვა სიტყვებით რომ ვთქვათ, რა ზღუდავს ფოტოსინთეზს სხვადასხვა მცენარეთა სხვადასხვა გარემოში? ამ ორ კითხვაზე პასუხი წარმოადგენს მცენარეთა პროდუქტიულობის გაზრდას. ამ ორ კითხვაზე პასუხის მიღება უაღრესად რთულია. ასზე მეტი ცილა მონაწილეობს ფოტოსინთეზში. მაშინაც კი, თუ მანიპულირება შემოიფარგლება მხოლოდ ცილის ოდენობის შეცვლით, ვიდრე მისი თვისებები, პოტენციური ტრანსპოზიცია მილიონებში გადავა.
იმის გათვალისწინებით, რომ მრავალჯერადი ცილის საჭიროება ერთდროულად უნდა იყოს ინჟინერირებული, აშკარა ხდება, რომ თითოეული ცილის ოდენობის ექსპერიმენტულად და რეგულირება ან შემცირება და მისი გავლენის შემოწმება ნაყოფიერებაზე არაეფექტურია. ამ პრობლემის გადასაჭრელად ერთი პოტენციური გზა არის სისტემური მიდგომა. სისტემური მიდგომაა მათემატიკური მოდელების შემუშავება, რომელთა საშუალებითაც შესაძლებელია სარწმუნოად სიმულაცია მოახდინოს ფოტოსინთეზში ჩართული ბიოფიზიკური და ბიოქიმიური პროცესების შემუშავება, შემდეგ კი ჩატარდეს რიცხვითი ექსპერიმენტები, რათა დადგინდეს სამიზნეები ინჟინერზე უფრო მაღალი პროდუქტიულობისთვის.
მე გამოვიყენებ ფოტოსინთეზური ნახშირბადის მეტაბოლიზმის მოდელს, რომ განვაზოგადო სისტემების შემუშავების მოდელების ძირითადი მეთოდოლოგია და ევოლუციური ალგორითმი გამოვიყენო მეტაბოლიზმში აზოტის გამოყოფის ოპტიმალური განაწილება, CO2-ის მოპოვების უფრო მაღალი ფოტოსინთეზური მაჩვენებლისთვის. ასევე მოკლედ აღვწერ ფოტოსინთეზის სისტემური მოდელების სხვა მნიშვნელოვან გამოყენებებს, რომლებიც მოიცავს გენომიური მონაცემების ფიზიოლოგიურ მონაცემებთან დაკავშირებას, ფოტოსინთეზში დინამიური სიგნალების შესწავლას, ფოტოსინთეზურ აპარატში ცვლილებების მნიშვნელობის შესწავლას ფოტოსინთეზური ეფექტურობის ასამაღლებლად და მუშაობის საფუძველს. ეკოფიზიოლოგიური მოდელები.
მცენარეთა ბიოლოგიის კვლევის ერთ-ერთი მთავარი სამიზნეა ფოტოსინთეზის გაზრდა
ბოლო ოთხი ათწლეულის განმავლობაში, მცენარეთა მეცხოველეობამ მნიშვნელოვანი ზრდა გამოიწვია მრავალი მოსავლის პოტენციურ მოსავალზე. პოტენციური მოსავლიანობა განისაზღვრება, როგორც მოსავლიანობა, რომლის მიღწევაც გენოტიპს შეუძლია მიაღწიოს ბიოტიკური და აბიოტიკური სტრესის გარეშე. მონტეიტი (1977) პრინციპების დაცვით, მოცემულ ადგილას მოსავლისა და პირველადი წარმოების (Y) პოტენციური მოსავალი (Pn) განისაზღვრება შემდეგით:
Pn = St.εi. εc/k …. (1a)
Y = η.Pn …. (1b)
სად St ინციდენტის მზის სხივების ყოველწლიური ინტეგრალია (მ.ჯ. m-2), εi ეფექტურობა, რომლითაც გამოსხივება ხვდება მოსავალს; εc ეფექტურობა, რომლითაც ხვდება გამოსხივება გარდაქმნილ ბიომასაში; η მოსავლის ინდექსი ან ეფექტიანობა, რომლითაც ბიომასის დაყოფა ხდება მოსავალ პროდუქტად; და k მცენარეთა მასის ენერგიის შემცველობა (მ.ჯ. g-1). St განისაზღვრება საიტის მიხედვით და წელი. k ძალიან განსხვავდება მცენარეულ ორგანოებს შორის, საშუალოდ საშუალოდ 17 მჯ/კგ, თუმცა მარცვლეულის მნიშვნელოვანი ნავთობის შემცველობა შეიძლება ჰქონდეს უფრო მაღალი ენერგიის შემცველობას (რობერტსი და სხვ., 1993). Pn არის პირველადი პროდუქტიულობა, ანუ მცენარეული ბიომასის მთლიანი წარმოება მზარდი სეზონის განმავლობაში. განტოლების საფუძველზე 1, Y განისაზღვრება კომბინირებული პროდუქტით სამი ეფექტურობა, თითოეული აღწერს მოსავლის ფართო ფიზიოლოგიურ თვისებებს: εi, εc, და η. ε1 განისაზღვრება ტილოების განვითარებისა და დახურვის სიჩქარე და ტილოების ხანგრძლივობა, ზომა და არქიტექტურა. εc არის ტოპიოპედიაში შემავალი ყველა ფოთლების ერთობლივი ფოტოსინთეზური განაკვეთის ფუნქცია, ნაკლებია მოსავლის რესპირატორული დანაკარგები (გრძელი და სხვ., 2006).
განტოლების 1 კონტექსტის გათვალისწინებით, ბოლო 40 წლის განმავლობაში პოტენციური მოსავლიანობის მატებამ ძირითადად განაპირობა ჰჰ και εi– ის ზრდა, ხოლო რეალიზებული მოსავლიანობა გაუმჯობესდა უკეთესი განაყოფიერებისა და დაავადებებისგან დაცვის გაუმჯობესების გზით, თავის მხრივ, გაიზარდა εi (მიმოხილულია: გრძელი და ალ., 2006). როგორ შეგვიძლია კიდევ უფრო გავზარდოთ Y? თანამედროვე ჯიშების ჯანსაღი კულტურები ოპტიმიზებულ დაშორებით დაშორებით ახდენენ ხელმისაწვდომი გამოსხივების უმეტესობას მათი მზარდი სეზონის განმავლობაში, რაც ზღუდავს εi გაუმჯობესების პერსპექტივებს. მარცვლეულის თანამედროვე ჯიშებში მარცვლეული შეიძლება წარმოადგენდეს მთლიანი ბიომასის 60% მოსავალს (ევან, 1993). მარცვლეულის მოსავლის ინდექსი, როგორც ჩანს, უახლოეს ზღვარს უახლოვდება, იმის გათვალისწინებით, რომ მცენარის სხეულში უნდა დარჩეს ბიომასის მინიმალური რაოდენობა, იმის უზრუნველსაყოფად, რომ სასიცოცხლო საკვები ნივთიერებები და მარაგები შეიძლება გადავიტანოთ მარცვლეულში, და გავითვალისწინოთ უჯრედის კედლის მასალები, რომლებიც არ შეიძლება. გადაგვარებულია. ბიომასის კულტურების მოსავლის ინდექსი კიდევ უფრო მაღალია. თუ η და εi ახლოვდება ზედა ზღვარს, პოტენციური მოსავლიანობის შემდგომი ზრდა მიიღწევა მხოლოდ εc– ის ზრდით, რაც განისაზღვრება ფოტოსინთეზით და სუნთქვით.
მაშ, რა არის თეორიული მაქსიმალური εc, და რა არის რეალიზებული εc? თეორიული ანალიზით ვარაუდობენ, რომ მაქსიმალური εc ამისთვის C3 და C4 არის 4.6% და 6%, შესაბამისად (სურათი 1, იხ. გაანგარიშების დეტალები იხ. ჟუ და სხვ., 2008).
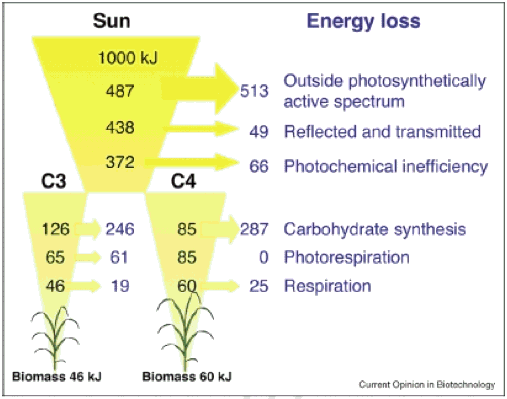
სურათი 1. ენერგიის მინიმალური დანაკარგები, რომელიც გამოითვლება 1000 კჯ ინციდენტის მზის სხივში, მცენარის ფოტოსინთეზური პროცესის თითოეულ დისკრეტულ ეტაპზე, რადიაციის შეწყვეტიდან, ბიომასის შემადგენლობაში შენახული ქიმიური ენერგიის ფორმირებამდე. განიხილება როგორც C3 და C4 (NADP-Malic ფერმენტის ტიპი) ფოტოსინთეზი. გამოთვლებით ვარაუდობენ ფოთლის ტემპერატურას 30 ° C და ატმოსფერული [CO2] 380 გვ. ისრები მიუთითებენ ენერგიის დანაკარგების ოდენობაზე სხვადასხვა პროცესებისთვის (ჟუ და სხვ., 2008).
C3 კულტურებისთვის ყველაზე მაღალი მოკლევადიანი ეფექტურობაა დაახლოებით 0,035, ხოლო C4– სთვის დაახლოებით 0,043; სფეროში მიღწერილი εc ამ მაქსიმალური მნიშვნელობების მხოლოდ 1/3-ს შეადგენს, რაც მიგვითითებს იმაზე, რომ არის მნიშვნელოვანი ადგილი, რომლითაც შესაძლებელი გახდება ε – ების კიდევ უფრო გაზრდა (მიმოხილულია ჟუ და სხვ., 2008). პირდაპირი მტკიცებულება იქნა მოწოდებული მცენარეთა მიერ, რომლებიც გაიზარდა Free Air CO2 გამდიდრებით, სადაც იზრდება როგორც ფოტოსინთეზური სიჩქარე, ასევე მცენარეთა პროდუქტიულობა (გრძელი და სხვები, 2004).
როგორ შეგვიძლია განვსაზღვროთ მაღალი პროდუქტიულობისთვის ინჟინერიის ახალი გზები? როგორც ადრე განვიხილეთ, ექსპერიმენტული მიდგომები ხდება არაეფექტური ახალი საინჟინრო მიზნების დადგენაში. აქ მე გამოვიყენებ ფოტოსინთეზური ნახშირბადის მეტაბოლიზმის მოდელს, რომ განვაზოგადო სისტემური მიდგომა, რათა დადგინდეს ახალი მიზნები, რაც უფრო მაღალია ფოტოსინთეზური ენერგიის გადაქცევის ეფექტურობის შესაქმნელად.
სისტემების ბიოლოგიის მიდგომა ინჟინერიისთვის ახალი შესაძლებლობების დადგენის მიზნით, უფრო მაღალი პროდუქტიულობისთვის
ამ ნაწილში მოკლედ აღვწერ სისტემური მოდელის შემუშავების ძირითად პროცედურას და შემდეგ აღვწერ სისტემების მოდელის ევოლუციურ ალგორითმებთან შერწყმის მეთოდს, რათა აღვნიშნოთ ინჟინერზე ახალი მიზნების მისაღწევად უმაღლესი პროდუქტიულობისთვის. ამ მონაკვეთის ბოლოს, მე განვმარტავ, თუ რატომ არ შეარჩია ევოლუცია ოპტიმალურ ფოტოსინთეზურ აპარატზე.
i) სისტემური მოდელის შემუშავების ძირითადი პროცედურა
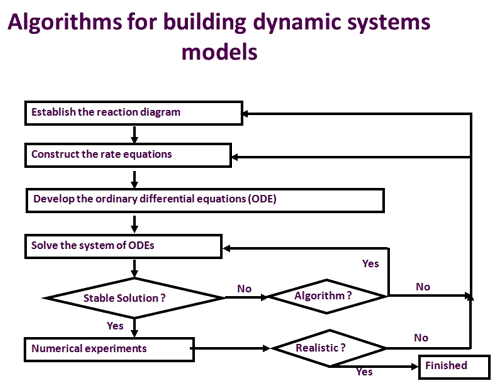
სურათი 2. ზოგადი ალგორითმი დინამიური სისტემების მოდელის შესაქმნელად. სისტემური მოდელის შემუშავებაში ჩართული პროცესი მოიცავს რეაქციის დიაგრამის დამკვიდრებას, განაკვეთის განტოლებების მშენებლობას, ჩვეულებრივი დიფერენციალური განტოლების მშენებლობას, მოდელის შედეგების ტესტირებასა და დადასტურებას ექსპერიმენტული დაკვირვებებით. ეს პროცესი იმეორებს მანამ, სანამ მოდელს არ შეუძლია ერთგულად მოახდინოს რეაგირება გენეტიკური ან გარემოსდაცვითი დარღვევების შესახებ.
გრაფიკი 2 ასახავს მეტაბოლური პროცესის კინეტიკური მოდელის შემუშავების პროცესს. ჯერ ერთი, ამ პროცესში მჭიდროდ ჩართული რეაქციები უნდა იყოს შედგენილი, რათა შეიქმნას რეაქციის დიაგრამა. შემდეგ, შემუშავებულია დიაგრამაში ჩართული თითოეული რეაქციისათვის განაკვეთის განტოლება. სისტემური მოდელი, რომელიც წარმოადგენს ამ მეტაბოლურ პროცესს, ხდება ჩვეულებრივი დიფერენციალური განტოლების (ODE) შემუშავებით პროცესში, რომელიც მონაწილეობს თითოეულ მეტაბოლიტზე. თითოეული ODE აღწერს მეტაბოლიტის კონცენტრაციის ცვლილების სიჩქარეს დროზე, ფორმირების ტემპებით მინუს ამ მეტაბოლიტის მოხმარების სიჩქარეს. მაგალითად, ფოტოსინთეზური ნახშირბადის მეტაბოლიზმში RuBP (რიბულოზა-1,5-ბისფოსფატი) იქმნება Ru5P (რიბულოზა-5-ფოსფატი) ფოსფორილირების გზით რეაქციის საშუალებით:
Ru5P+ATP→ ADP + RuBP …. 2
ის მოიხმარენ როგორც RuBP კარბოქსილაციას, ასევე RuBP ჟანგბადით:
RuBP + CO2 → 2 PGA …. 3
RuBP + O2 → PGA + PGCA …. 4
სადაც PGA არის 3-ფოსფოგლიცერატი და PGCA არის 2-ფოსფოგლიკოლატი. თუ ვივარაუდებთ, რომ RuBP ფოსფორილირების ტემპი არის vrp, ხოლო RuBP კარბოქსილაციის (განტოლება 3) და ჟანგბადის (განტოლება 4) განაკვეთები არის vc და vo, შესაბამისად, მაშინ RuBP კონცენტრაციის ტემპი დროით გამოიხატება:
d[RuBP]/dt = vrp-vc-vo …. 5
ყველა ODE ერთობლივად ქმნის ჩვეულებრივი დიფერენციალური განტოლების სისტემას, რომლის გადაწყვეტა შეესაბამება მეტაბოლური პროცესის სხვადასხვა ფიზიოლოგიურ და დინამიურ მდგომარეობას.
მას შემდეგ, რაც შეიქმნა ჩვეულებრივი დიფერენციალური განტოლებათა სისტემა, საჭიროა ციფრული ექსპერიმენტების სერია მოდელის შესამოწმებლად და გადამოწმებაზე. ჩვეულებრივ, სისტემურმა მოდელმა უნდა აჩვენოს სტაბილური მდგომარეობის მოპოვების უნარი, ძლიერი დარღვევა გარე არეულობისგან და ჩვეულებრივ დაკვირვებული მოვლენების პროგნოზირების უნარი. მაგალითად, ფოტოსინთეზური ნახშირბადის მეტაბოლიზმის მოდელმა მიაღწია სტაბილურ მდგომარეობას დაახლოებით 200 წამში (ჟუ და სხვ. 2007). იგი ერთგულად ახდენდა ჩვეულებრივ გაზომვას A-Ci მრუდი, ე.ი. CO2 ათვისება ინტერსელექტრულით CO2 კონცენტრაცია (Ci) მრუდი, ნორმალური (21%) და დაბალი (2%) ჟანგბადის დონე. გარდა ამისა, მკვეთრი შეშუპების შემდეგ ინტერცელულარული O2 კონცენტრაცია, მოდელს შეუძლია სწრაფად დაიბრუნოს ახალი სტაბილური მდგომარეობა ფოტოსინთეზის რეალისტური მაჩვენებლით. დაბოლოს, მოდელმა ერთგულად შეადარა ფოსფატს შეზღუდული ფოტოსინთეზი, ტრიოზის ფოსფატის გამოყენების დაბალი სიჩქარის პირობებში (ჟუ და სხვ., 2007). ამ დადასტურების შემდეგ, ფოტოსინთეზური ნახშირბადის მეტაბოლიზმის მოდელი შეიძლება გამოყენებულ იქნას ახალი მიზნების იდენტიფიცირებისთვის, უფრო მაღალი ფოტოსინთეზური ეფექტურობის შესაქმნელად.
ii) სისტემების მოდელის და ევოლუციური ალგორითმის შერწყმა ინჟინერზე ახალი მიზნების მისაღწევად მაღალი პროდუქტიულობისთვის
ტრადიციულ მეცხოველეობაში, სხვადასხვა მცენარეებს შორის ჯვრები ქმნის ცვალებადობას გარკვეულ თვისებებში, მაგალითად, მოსავლის მოსავლიანობას. თესლები მათთვის, ვისაც აქვს უმაღლესი მახასიათებლის თვისებები, შეირჩევა და გამოიყენება შემდეგი თაობისთვის ინდივიდების შესაქმნელად. მრავალი თაობის შემდეგ, შერჩეული თვისებები თანდათან იხვეწება ამ პროცესის განმავლობაში. ამ მეთოდმა განაპირობა მოსავლის პროდუქტიულობის მკვეთრი ზრდა და შექმნა „მწვანე რევოლუცია“. ტრადიციული მეცხოველეობის დროს თაობებზე უკეთესი თვისებების შერჩევის ანალოგიურია, სისტემების მოდელების ევოლუციური ალგორითმის შერწყმა საშუალებას იძლევა ფოტოსინთეზური აპარატის სიმულაცია მოახდინოს უფრო მაღალი ფოტოსინთეზური მაჩვენებლისთვის.
ევოლუციური ალგორითმი არსებითად ასახავს ბუნებრივი გადარჩევის პროცესს. რეალურ ბუნებრივ შერჩევაში, შერჩევის წნევა არის გადარჩენის და უფრო მაღალი ფეკალუტის უზრუნველყოფა. ევოლუციური ალგორითმის მიერ გამოყენებული ხელოვნური შერჩევისას, შერჩევის წნევა უნდა ჰქონდეს უფრო მაღალი ფოტოსინთეზური CO2-ის დაქვეითების მაჩვენებელი, იგივე რაოდენობით აზოტის ინვესტიცია, ფერმენტებში ნახშირბადის მეტაბოლიზმის ფერმენტებში. გარდა ამისა, თითოეულ თაობას აქვს მრავალი ინდივიდი, რომელთაგან თითოეული წარმოადგენს ფერმენტული კონცენტრაციების ერთ ჯგუფს, რომლებიც საჭიროა ნახშირბადის მეტაბოლიზმის მოდელის შესასრულებლად. თითოეული ინდივიდის, ან ფერმენტის კონცენტრაციის ერთობლიობისთვის, არსებობს CO2–ის შეფერხების ერთი სტადიური მდგომარეობა. თითოეულ თაობაში, CO2-ის უფრო მაღალი ფოტოსინთეზური შემცველობის მქონე ინდივიდი შეირჩევა მომავალი თაობის “თესლისთვის”. მომდევნო თაობებისთვის ინდივიდების შესაქმნელად, შერჩეული ინდივიდი დუბლირებული იქნება მოსახლეობის გენერირებისთვის, მაშინ ამ პოპულაციაში ინდივიდები “მუტაციდებიან” (ე.ი. ფერმენტის კონცენტრაციებზე შემთხვევითი ვარიაციების დამატება). შერჩევისა და მუტაციის ეს პროცესი განმეორდება, ხოლო CO2–ის ფოტოსინთეზური შემცველობის მაჩვენებელი თანდათანობით მატულობს თაობებში. ეს პროცესი მთავრდება, როდესაც ფოტოსინთეზის სიჩქარე არ შეიძლება გაიზარდოს უფრო თაობებთან ერთად (ჟუ და სხვ., 2007). აზოტის საბოლოო განაწილება ფერმენტის ნაკრებში განიხილება, როგორც აზოტის ოპტიმალური განაწილება (სურათი 3).
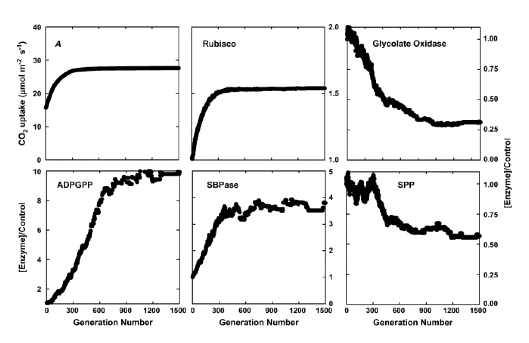
დიაგრამა 3. ნახშირბადის მეტაბოლიზმის ფერმენტებს შორის რესურსების ოპტიმალური დაყოფის შერჩევა, სინათლის გაჯერებული ფოტოსინთეზური სიჩქარის მაქსიმალურად მისაღწევად. A, მსუბუქი გაჯერებული ფოტოსინთეზური მაჩვენებლის გაზრდა 1.500 თაობის არჩევაში, ევოლუციური ალგორითმის გამოყენებით. B to F–ს აჩვენებს ამ ფერმენტების ხუთეულში ოდენობის ცვლილებებს მათი საწყისი მნიშვნელობების მიმართ ამ 1,500 თაობის განმავლობაში. შერჩევა მიმდინარეობდა მსუბუქი გაჯერებით, ხოლო Ci – 280 μმოლი მოლი-1.
აბრევიატურა:
A: ფოტოსინთეზური CO2-ის მიღების მაჩვენებელი
ADPGPP: ADP გლუკოზის პიროფოსფორილიზა
SBPase: სედოჰეპულოზა-l,7-ბისფოსფატაზა
SPP: საქაროზა ფოსფატის ფოსფატაზა
FBPase: ფრუქტოზა-2,6-ბისფოსფატაზა.
ჟუ და სხვ. (2007).
აზოტის ოპტიმალური განაწილების გამოვლენის შემდეგ, ჩვენ შეგვიძლია გამოვიყენოთ ის, რომ გამოვთვალოთ ზრდის მოთხოვნა თითოეული ფერმენტისათვის,
Demand[i] = [Enzyme[i]_opt]/ [Enzyme[i]_initial] …. (6)
სადაც [Enzyme[i]_opt] წარმოადგენს აზოტის ოპტიმალურ კონცენტრაციას და [Enzyme[i]_initial] წარმოადგენს აზოტის საწყის კონცენტრაციას. თუ ფერმენტზე მოთხოვნა 1-ზე მეტია, ფერმენტული აქტივობა უნდა გაიზარდოს; წინააღმდეგ შემთხვევაში, ფერმენტის აქტივობა უნდა დაიწიოს აზოტის გასათავისუფლებლად სხვა ფერმენტების სინთეზისთვის, რათა მიიღოთ CO2 მაქსიმალური მიღება, რაც მიიღებს ოპტიმალურ რაოდენობას (ჟუ და სხვ., 2007). ამ მიდგომის გამოყენებით, ჩვენ დავანახვეთ, რომ რუბისკო, SBPase და FBPase უნდა გაიზარდოს უფრო მაღალი ფოტოსინთეზური CO2 მაჩვენებლისთვის (ჟუ და სხვ., 2007). გარდა ამისა, ჩაძირვის ტევადობის ზრდა, ანუ ტრიოოს ფოსფატის (PGA, ან დიჰიდროქსიაცეტონის ფოსფატის, ან გლიცერალდეჰიდის 3-ფოსფატის) მოხმარების შესაძლებლობა, სახამებლის, საქაროზის ან სხვა მაკრომოლეკულების სინთეზის საშუალებით. ამ ფერმენტების საქმიანობის გასაუმჯობესებლად საჭირო აზოტი სავარაუდოა, რომ გამოიყოფა ფოტო – რესპირატორული გზიდან (ჟუ და სხვ., 2007). SBPase, ერთ – ერთი ფერმენტი, რომელიც გაიზარდა, რომ გაიზარდოს უფრო მაღალი ფოტოსინთეზის მოსაპოვებლად, თამბაქოზე მეტი გამოითქვა, რამაც გამოიწვია უფრო მაღალი ფოტოსინთეზის და ბიომასის წარმოება (ლეფებვარი და სხვები 2005 პოტენციური საინჟინრო მიზნები უმაღლესი ფოტოსინთეზისთვის.
ინჟინრისთვის მიზნების იდენტიფიცირების კიდევ ერთი მიდგომა არის მეტაბოლური კონტროლის ანალიზი (MCA) ნაკადის კონტროლის კოეფიციენტის გამოანგარიშებით (დაეცა 1997). MCA და ევოლუციური ალგორითმი ინჟინერულ მიზნებს წარმოადგენენ; ამასთან, ამ ორ მიდგომას შორის ფუნდამენტური განსხვავებებია. ფერმენტის ნაკადის კონტროლის კოეფიციენტი წარმოადგენს ამ ფერმენტის კონტროლს გარკვეულ ნაკადადზე, თუ ვიღებთ ყველა სხვა ფერმენტის დონის შეცვლას, ანუ, ეს კოეფიციენტი წარმოადგენს სისტემურ ნაკადადზე ამ ფერმენტის კონტროლის ადგილობრივ ხედვას. თავის მხრივ, ევოლუციური ალგორითმი განსაზღვრავს ყველა ფერმენტის ოპტიმალურ განაწილებას მეტაბოლიზმში, ერთდროულად შეცვლის ყველა ფერმენტის კონცენტრაციას. აქედან გამომდინარე, თეორიულად, ევოლუციურ ალგორითმს შეუძლია უფრო მაღალი ფოტოსინთეზის მიზნების იდენტიფიცირება სისტემის უფრო გლობალური თვალსაზრისით.
iii) რატომ არ მიიღეს ფოტოსინთეზი, მილიონობით წლის ევოლუციის შემდეგ, არ ვითარდება ოპტიმალური ფოტოსინთეზური მაჩვენებელი?
ევოლუციური ალგორითმის კომბინაცია სისტემურ მოდელებთან უზრუნველყოფს შესაძლო და ეფექტურ მექანიზმს ოპტიმალური ფოტოსინთეზური აპარატის იდენტიფიცირებისთვის. შეიძლება დაისვას ერთი ფილოსოფიური კითხვა: მილიონობით წლის ევოლუციის შემდეგ, რატომ არ განვითარდა მცენარეები ოპტიმალურ ფოტოსინთეზურ აპარატად? არსებობს რამდენიმე შესაძლო მიზეზი. ჯერ ერთი, ევოლუცია ირჩევს გადარჩენას და ფეტვიურობას, და არა პროდუქტიულობას. სტრესულ პირობებში გადარჩენის უზრუნველსაყოფად, გარკვეული რაოდენობის რესურსი, მაგალითად აზოტი, ინვესტიციდება დამცავ მექანიზმებში ფოტოსინთეზური ეფექტურობის ხარჯზე. მაგალითად, ჩვენ დავამტკიცეთ, რომ აზოტი ზედმეტი ინვესტიციით ხორციელდება ფოტო – რესპირატორული გზით (ჟუ და სხვ., 2007). ამასთან, ეს ინვესტიცია გადამწყვეტი მნიშვნელობა ენიჭება მცენარეთა გადარჩენას ძალიან ცხელ პირობებში. მიუხედავად იმისა, რომ ასეთი პირობები იშვიათია, შესაძლოა ისინი რამდენიმე ასეულ წელიწადში ერთხელ მოხდეს, ისინი შეიძლება საკმარისად ხშირი გახდნენ მცენარეების სრულად მოსაშორებლად, ასეთი დამცავი მექანიზმების გარეშე (ჟუ და სხვ., 2007) მეორეც, გლობალური კლიმატის ცვლილება კიდევ ერთი ფაქტორია. ატმოსფერული ატმოსფერული CO2 კონცენტრაცია ბოლო 25 მილიონი წლის განმავლობაში იყო დაახლოებით 280 გვ (ბარნოლა და სხვ., 1999), ინდუსტრიული რევოლუციის შემდეგ CO2-ის კონცენტრაციის სწრაფ ზრდამდე. მიმდინარე ფოტოსინთეზური ნახშირბადის მეტაბოლიზმი, მაგ., უფრო მაღალი ინვესტიცია ფოტომასტიური სისტემის გზაზე, შეიძლება წარმოადგენდეს აზოტის ოპტიმალურ გამოყოფას წინამორბედი რევოლუციის CO2 დონის (ჟუ და სხვ., 2007). ჩვენ ადრე დავანახეთ, რომ ფოტოსინთეზის ერთ – ერთი კრიტიკული ფერმენტი, რუბიკო, ნამდვილად არის ოპტიმიზირებული CO2–ის უფრო მაღალი მიღებისთვის 280 გვ ატმოსფერული CO2 კონცენტრაციისთვის (ჟუ და სხვ., 2004). კიდევ ორი დახვეწილი მიზეზი შეიძლება იყოს პასუხისმგებელი აზოტის ამჟამინდელი არა ოპტიმალური გამოყოფისთვის ფოტოსინთეზური ნახშირბადის მეტაბოლიზმში. SBPase-ის ქვე-ოპტიმალური ინვესტიცია შეიძლება დაკავშირებული იყოს SBPase–ის, როგორც მაკონტროლებელი წერტილის როლზე, ფოტოსინთეზური ნახშირბადის მეტაბოლიზმში; ხოლო ფერმენტების ზედმეტი ინვესტიცია ფოტომასპინძურ გზაზე შეიძლება ასევე გამოწვეული იყოს RuBP კარბოქსილაციასა და ჟანგბადობას შორის მკაცრი თანაფარდობით მოცემული რუბიკოს სპეციფიკისა და ატმოსფერული პირობების გამო (ჟუ და სხვ., 2007).
ფოტოსინთეზის სისტემების მოდელების სხვა ძირითადი პროგრამები
დინამიური სისტემების მოდელები ინტეგრირებისთვის ახალი მიდგომების დადგენის გარდა, სხვა პროდუქტებს იყენებენ, უფრო მაღალი პროდუქტიულობისთვის. მოიცავს:
ჯერ ერთი, სისტემების მოდელები იძლევა გენომიური მონაცემების პირდაპირ კავშირს ფენოტიპურ მონაცემებთან. სისტემების მოდელის მონაცემები არის ფერმენტის მოქმედებები ან ცილის შემცველობა, რომელთა მიღება შეგიძლიათ ან ლიტერატურის კვლევის ან პროტეომიკის საშუალებით. სისტემების მოდელების შედეგები არის მეტაბოლიტის კონცენტრაცია, ნაკადები და ფიზიოლოგიური პარამეტრები.
მეორე, სისტემების მოდელი შეიძლება გამოყენებულ იქნას ფოტოსინთეზური აპარატის მოლეკულური ცვლილებების ადაპტაციული მნიშვნელობის შესასწავლად ფოტოსინთეზურ ეფექტურობაზე. აქ ფოტოსინთეზური აპარატი განისაზღვრება, როგორც სხვადასხვა კომპონენტები (ფერმენტები, ელექტრონის გადაცემის მატარებლები და ა.შ.) და სხვადასხვა კომპონენტებს შორის ურთიერთქმედება ფოტოსინთეზში. როგორც უკვე აღვნიშნეთ, ევოლუციური ან გენეტიკური ალგორითმების გაერთიანება სისტემურ მოდელთან უზრუნველყოფს ეფექტურ საშუალებას, განსაზღვრული პირობებით ოპტიმალური ფოტოსინთეზური აპარატის იდენტიფიცირება. ეს განსხვავება ოპტიმალურ ფოტოსინთეზურ აპარატსა და არსებულ ფოტოსინთეზურ აპარატს შორის საშუალებას გვაძლევს, ინჟინერი შეიძინოთ უფრო მაღალი პროდუქტიულობისთვის. ეს განსხვავება ასევე მიგვითითებს ფოტოსინთეზური აპარატის სხვადასხვა თვისებების ადაპტაციის მნიშვნელობაზე.
მესამე, დინამიური სისტემების მოდელები შეიძლება გამოყენებულ იქნას ჰიპოთეზის შესამოწმებლად, ფოტოსინთეზთან დაკავშირებული დინამიური სიგნალების დამყარებული მექანიზმების შესახებ. ჩვენ გამოვიყენეთ ფოტოსისტემის II-ის გარშემო არსებული ძირითადი მოვლენების სისტემური მოდელი, რათა შევისწავლოთ ფლუორესცენტული ინდუქციის მრუდი მექანიკური საფუძველი, რომელიც ჩაწერილი იყო გაჯერებული მსუბუქი პულსის ანათების შემდეგ (ე.ი. მოკლე მსუბუქი პულსი (<1s), რომელსაც შეუძლია დახურვა ყველა ფოტოსინთეზის II რეაქციის ცენტრში) მუქი ადაპტირებული ფოთლები (ჟუ და სხვ., 2005).
მეოთხე, ფოტოსინთეზის სისტემური მოდელის გამოყენება შესაძლებელია ეკოფიზიოლოგიური მოსავლის სიმულაციის მოდელებში, რათა შესაძლებელი გახდეს პირდაპირი კავშირი გენომიდან ეკოსისტემაში. ინტეგრირებული მიკროსქემის (IC) კომპიუტერულმა დიზაინმა გადამწყვეტი როლი ითამაშა ახალი ახალი ტექნოლოგიების სწრაფ განვითარებაში. მოსალოდნელია, რომ მოდელს, რომელსაც შეუძლია გენომიდან ეკოსისტემასთან პირდაპირ დაკავშირება, შეუძლია იგივე როლი შეასრულოს. მიუხედავად იმისა, რომ სოფლის მეურნეობის ეკოსისტემების მოდელები შემუშავებულია გასული ნახევარი საუკუნის განმავლობაში, მათ ჯერ კიდევ არ შეუძლიათ უშუალოდ განსაზღვრონ მოსავლის მოქმედება მოსავლის გენომიდან და მისი ზრდის გარემოდან (Jones et al., 2003), თუმცა QTL- ების გამოყენებისთვის ბოლო რამდენიმე ძალისხმევა არ არსებობს. დნმ-სთან, რომლებიც მჭიდრო კავშირშია იმ გენებთან, რომლებიც ხაზს უსვამენ ხაზს) აგრო-ეკოსისტემის მოდელებში. (იინი et al., 2004).
ინტეგრირებული მოდელის შემუშავება, რომელსაც შეუძლია გენომი დაუკავშიროს ეკოსისტემას, ძალზე რთულია, რადგან ასეთ მოდელს არა მხოლოდ მექანიკურად უნდა აღინიშნოს პროცესები სხვადასხვა ორგანიზაციულ დონეზე, დაწყებული ეკოსისტემის დონიდან, კანაფის დონის, ფოთლებისა და უჯრედული დონის პროცესებზე, არამედ გააჩნია გენის გამოხატვის ცვლილებების აღწერა სხვადასხვა პირობებში და განვითარების სხვადასხვა ეტაპზე. ეჭვგარეშეა, რომ ასეთი მოდელის შემუშავება დასჭირდება მრავალი წლის განმავლობაში ძალისხმევას მთელი მცენარეთა სამოდელო საზოგადოების მიერ. იმის გათვალისწინებით, რომ ფოტოსინთეზი მცენარეთა ზრდისა და განვითარებისთვის მატერიალური და ენერგიის საფუძველია, ფოტოსინთეზის დეტალური მექანიკური მოდელი წარმოადგენს ასეთი ინტეგრირებული მოდელის კრიტიკულ კომპონენტს. ფოტოსინთეზური ნახშირბადის მეტაბოლიზმის მოდელის ინტეგრაცია აგრო-ეკოსისტემის მოდელებთან, შესაძლოა, პირველი ნაბიჯი იყოს ასეთი ინტეგრირებული გენომი-ეკოსისტემური მოდელის რეალიზაციისა.
ლიტერატურა:
ბარნოლა, ჯმ, რეინოს, დ ლორიუსი, გ. და ბარკოვი, ნი ისტორიული CO2, საწყისი ვოსტოკის ყინულის ბირთვი. ტენდენციები: კომპონენტი მონაცემების გლობალური ცვლილება. ნახშირორჟანგი ინფორმაციის ანალიზის ცენტრი, მუხის ქედი ეროვნული ლაბორატორია, ენერგეტიკის დეპარტამენტის, მუხის ქედი, ტნ, ამერიკის შეერთებული შტატები (1999).
ევანs ლტ მოსავლის ევოლუცია, ადაპტაციის და სარგებელი. CUP, კემბრიჯი. (1993)
დაეცა, დ.ა. გააზრება კონტროლის ცვლის. პორტლენდ პრესი, ლონდონი (1997).
ჯონსი, მაღალი ხე, მაღალიხე, გ, პორტერი, თ, კატარღა, ბსგ, ბაკალავრი, ჰანტი, ლ.ა., უილკინსი, პაუ, ლეო, უ, გიზმანი, აჯ, რიჩი, ჯტ DSSAT მოჭრა სისტემის მოდელი. ევროპული ჟურნალი აგრონომია 18, 235-265 (2003).
ლეფებვარი, ს., ლოუსონი, თ., ზახლენიუკი, ოვ, ლოიდ, ჯ.კ. რაინსი, კალიფორნია გაზრდილი სედოჰეპულოზა-1,7-ბისფოსფატაზა საქმიანობის ტრანსგენური თამბაქოს მცენარეთა უწყობს ფოტოსინთეზის და ზრდის ადრეულ ეტაპზე განვითარებას. მცენარეთა ფიზიოლი. 138, 451-460 (2005).
დიდხანს, ჟუ, გ., ნაიდუ, და ორტ, დრ შეუძლია გაუმჯობესება ფოტოსინთეზის ზრდა მოსავლის? მცენარეთა უჯრედი გარემო. 29, 315-330 (2006).
დიდხანს, აინსორთი ე., როჯერსი ა ადგილი ამოდის ატმოსფერული ნახშირორჟანგი: მცენარეთა წინაშე მათი მომავალი. წლიური მიმოხილვა მცენარეთა ბიოლოგია, 55, 591-628 (2004).
მონტეიტი, ჯლ კლიმატი და ეფექტურობის მოსავლის ბრიტანეთში. ფილო. ტრანს. რ სოკ. ლონდონში 281, 277-294 (1977).
რობერტსი მ.ჯ., დიდხანს ტიესენ ლლ და მძივი კლ საზომი მცენარეთა ბიომასის და წმინდა პირველადი წარმოების ბალახოვანი მცენარეულობა: ფოტოსინთეზი და წარმოების ცვალებად გარემოში: საველე და ლაბორატორიული სახელმძღვანელო (რედ, გააკეთე დარბაზი, ჯმო სკურლოკი, ბოლჰარი ნორდენკამფ, ლეეგუდი და გრძელი), გვ. 1-21. ჩეპმენი და ჰოლი, ლონდონი (1993).
იინი, სტრუიკი, კომპიუტერი და კოროფი, მ.ჯ. როლი მოსავლის ფიზიოლოგია პროგნოზირების გენი-ფენოტიპის ურთიერთობები. ტენდენციები მცენარეთა მეცნიერება. 9, 426-432 (2004).
ჟუ, გ, პორტისი კინგი, რ და დიდხანს, სურდა ტრანსფორმაციის C3 მოსავალი მცენარეთა უცხოელ რუბისკო ზრდა პროდუქტიულობა? კომპიუტერული ანალიზი ექსტრაპოლაციის ეხლა კინეტიკური თვისებები ტილო ფოტოსინთეზი. მცენარეთა უჯრედი გარემო. 27, 155-165 (2004).
ჟუ, გ კომპიუტერული მიდგომები სახელმძღვანელო ბიოტექნოლოგიური გაუმჯობესება მოსავლის ფოტოსინთეზური ეფექტურობა. დოქტორანტი თეზისი, ილინოისის უნივერსიტეტის. ურბანა, ილ. (2004).
ჟუ, გ, გოვინჯი, ბეიკერი, კაცი, დესლერი, ა., ადგილი, დრ, გრძელი, პ ქლოროფილი ფლუორესცენტი ინდუქციური კინეტიკა ფოთლები პროგნოზით, მოდელი აღწერს ყოველი დისკრეტული კომპლექტი აგზნების ენერგია და ელექტრონული გადაცემის უკავშირდება ფოტოსისტემით II. მცენარეთა 223, 114-133 (2005).
ჟუ, გ, დე შტურლი, ე. დიდხანს, ოპტიმიზაციის რესურსების გადანაწილების შორის ფერმენტების ნახშირბადის ცვლის შეიძლება მკვეთრად გაიზარდოს ფოტოსინთეზური განაკვეთი: რიცხვითი მოდელირება გამოყენებით ევოლუციური ალგორითმი. მცენარეთა ფიზიოლი. 145, 513-526 (2007).
ჟუ, გ,, გრძელი, და ადგილი, რა არის მაქსიმალური ეფექტურობა, რომელიც ფოტოსინთეზის შეუძლია გარდაქმნას მზის ენერგიის შევიდა ბიომასის? ტენდენციები ბიოტექნოლოგია 19, 153-159 (2008).
8/17/08